
|
||||||
| Answers | Species | Science | Community | Home | ||
|
From The Condor 93:279-285 The Effect of Floral Abundance on Feeder Censuses of Hummingbird PopulationsDavid W. Inouye William A. Calder Nickolas M. Waser
AbstractNumbers of Broad-tailed Hummingbirds (Selasphorous platycercus) captured each summer from 1979-1989 at the Rocky Mountain Biological Laboratory were quite variable, ranging from 115 (1981) to 348 (1989), with new birds usually outnumbering returning (previously banded) birds. Capture numbers were negatively correlated with the abundance of four species of flowers they visited, Erythronium grandiflorum, Delphinium nelsonii, Ipomopsis aggregata and Delphinium barbeyi; flower numbers were also highly variable during the study period. Since most of the captures were at feeders, these data suggest that in years with high floral abundance feeders are less attractive, while in years with low floral abundance hummingbirds with nests or territories at greater distances increase their use of the feeders. This interpretation is supported by seasonal variation in use of feeders, which is highest during the beginning and end of the season when floral abundance is lowest. Estimates of hummingbird density based on activity at feeders may thus be affected by the availability of floral food resources. Key words: Banding; census; floral abundance; Broad-tailed Hummingbird;
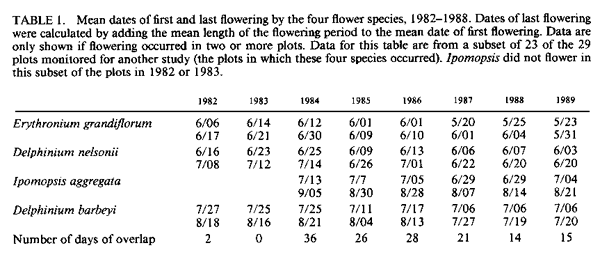
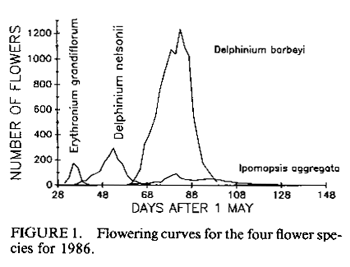
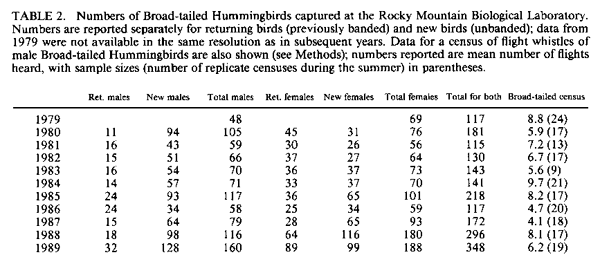
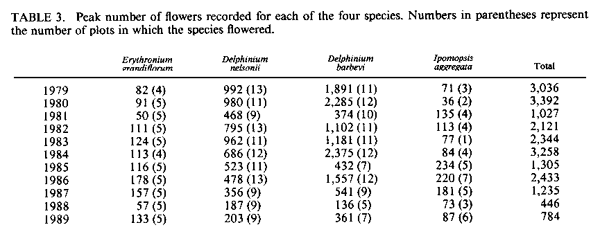
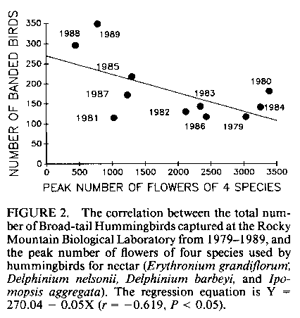
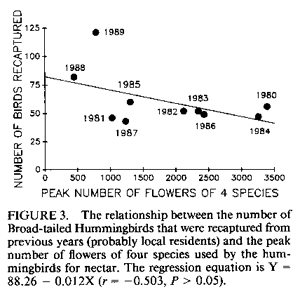
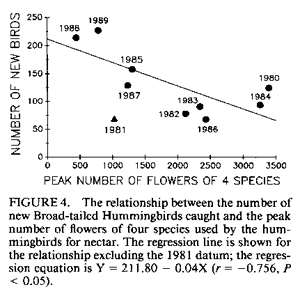
population size; resources. IntroductionMany bird populations vary in size from year to year (e.g., Mulvihill and Leberman 1987), or even within years. In some cases this variation can be ascribed to particular environmental or demographic variables. For example, droughts may affect reproduction and survivorship through an effect on food resources (Gibbs and Grant 1987, Grant and Grant 1989), cold temperatures may result in decreased winter survivorship, or small population sizes may result in local extinctions. However, variation in population size usually remains unexplained. Hummingbird numbers seem to vary considerably, as perceived by numbers at feeders. Apparent lows are popularly attributed to environmental catastrophes on the wintering grounds for many species in Central America, such as volcanic eruptions, forest fires, and the use of pesticides. While these perceptions lack the rigor of scientific methodology, casual observations by numerous individuals nevertheless suggest that these are plausible explanations. Accurate estimates of hummingbird population size would permit more careful consideration of the significance of this variation. A variety of techniques can be used to assess population size. Banding offers an advantage over techniques based on visual sightings, or counts of song or flight noise that may be conducted more easily, in that individuals are identified and counted only once. We have carried out long-term studies using two methods, banding and flight noise, on a resident population of Broad-tailed Hummingbirds (Selasphorus platycercus Swainson). During our 18-year study we have observed substantial year-to-year variability in abundance of Broad-tailed Hummingbirds in Colorado. There is also significant annual variation in the availability of floral resources used by these birds. Here we examine the relationship between the abundance of the hummingbirds, as indicated by banding studies, and floral resources. Materials and MethodsThis study was conducted at the Rocky Mountain Biological Laboratory (RMBL),Crested Butte, CO, at an elevation of approximately 2,900 m. Broad-tailed Hummingbirds, the only resident breeding species of hummingbird at RMBL, have been studied extensively at RMBL (e.g., Calder 1975, 1981, 1984, 1985; Calder and Booser 1973; Calder et al. 1983; Waser 1978, 1988; Waser and Inouye 1977; Waser and Price 1983). Hummingbirds were captured and banded each year at two to three different sites separated by about 500 m. Most birds were captured at feeders at cabins using cages, mist nets, or butterfly nets, with some additional mist net captures in meadows around RMBL. Although the banding program began in 1971, it was intensified beginning in 1979 to the point where we think that almost all resident birds were banded; by the end of 1979 and subsequent summers the continued netting and trapping yielded primarily recaptures of birds already recorded for that year. In addition to data from banding, a potential correlate of population size was also measured: the numbers of male Broad-tailed Hummingbirds heard during 10 min of listening from a fixed location in a meadow at RMBL at a standard time of day (noon). Such censuses were conducted every two to four days throughout each summer. Males of this species make a mechanical wing whistle that is audible up to 100 m away (Miller and Inouye 1983). Data on flower abundance were collected from 29 2 x 2-m plots, located within 500-1,000 m of banding sites. Approximately every other day for most of the growing season (early May to mid-September), all flowers in the plots were counted. Plots were scattered among habitats including a dry rocky meadow (7 plots), aspen forest (2 plots), wet meadow (9 plots), dry (but not rocky) meadow (3 plots), aspen-meadow interface (3 plots) and willow-meadow interface (S plots). For each species in every plot, the peak number of flowers produced during a given year was determined. For analyses in this study, the peak numbers for each species were added across plots for each year to produce a single value of peak number of flowers. Not all species occurred in all plots; for example, Ipomopsis was only found in 1-7 of the plots in any given year. However, representation of species in the plots approximated their abundance at the study site. Data from four species of flowers that are among those most commonly visited by hummingbirds at the study site were used for this analysis. They were Erythronium grandiflorum (glacier lily; Liliaceae), Delphinium nelsonii (Nelson's larkspur; Ranunculaceae), Delphinium barbeyi (tall larkspur; Ranunculaceae), and Ipomopsis aggregata (scarlet gilia; Polemoniaceae). Only the latter fits the classical characterization of a hummingbird-pollinated plant (long, red, tubular flowers). Data were not included for other flowers used by hummingbirds, such as Castilleja spp. (Scrophulariaceae), or Aquilegia elegantula (Ranunculaceae) because they were relatively rare. The four flower species used for this study span much of the flowering season (Table l; Fig. 1). Erythronium, with large, yellow, pendant lily flowers, is one of the first species to flower at the study site, and is also visited by bumblebees and other bees. The two blue-flowered Delphinium species are also visited frequently by bumblebees, while hummingbirds are the primary visitors to Ipomopsis (Waser 1982), although bumblebees commonly rob the nectar of Ipomopsis flowers in some years (Inouye 1980). Despite the variation among years in date of first flowering (Table 1), the relative sequence of flowering is generally maintained, and the arrival and nesting of the birds appear to be synchronized with flowering. ResultsTable 2 summarizes the numbers of Broad-tailed Hummingbirds captured at RMBL from 19791989. The sex ratio among captures was biased in favor of males in 1980 and females in 1979 and 1988 (binomial probability test, P < 0.05). The total number of birds captured ranged from a low of 115 in 1981 to a high of 348 in 1989. From 1980-1987 the number of recaptured (probably resident) birds was more constant than the number of newly captured (probably migrant) birds. The number of birds captured was highest in 1988 and 1989, when the numbers of flowers were lower than in any other year. These were the only two consecutive years of low flower numbers during the study, and this sequence was apparently responsible for the large numbers of new birds (214) banded in 1988, and birds recaptured in 1989; many of the recaptures were first banded in 1988. There was also a large number of new hummingbirds in 1985. Data from the census of wing-whistle noises are also shown in Table 2; there was no significant correlation between these census data and the numbers of banded males (r < 0.100, n = 11, P > 0.05), or between the number of flowers and the number of males heard in the census of flight noises (r = 0.142, P > 0.05). There was much variation among years in the number of flowers produced by each of the four plant species (Table 3). The reasons for this are not completely understood, although two factors appear to be important. First, there is a significant correlation between the number of flowers of both Delphinium species produced each summer and the amount of snowfall the previous winter (Inouye 1989); there is no such correlation for the other two species. Second, every four to five years (1976, 1981, 1985, 1989) since 1973 (when flower data were first collected from these plots) there have been late (mid-June) hard frosts (sometimes accompanied by snow) that have killed flower buds of many species (Inouye 1988). These two environmental events appear to be responsible for much of the annual variation in flower number in Delphinium species and for some of the other species in these meadows. There is a significant negative correlation between the number of birds captured and the peak number of flowers (Fig. 2; r = -0.619, P < 0.05) for the period 1979-1989. If the capture data are broken down into numbers of recaptured birds and new birds (Table 2; data only available since 1980) and then correlated with flower numbers, the negative correlation for recaptured birds is not significant (Fig. 3; r = -0.503, 0.1 > P > 0.05), while the correlation for new birds is significant only if the 1981 datum is excluded (Fig. 4; r = -0.756, P < 0.05). The year with the greatest deviation from the regression in Figure 4 is 1981, with fewer birds captured than predicted by the regression equation (and the lowest number of birds during the I l-year study). This year was also unusual in another regard: it was the only year in the past 15 years in which there was legitimate (not nectar robbing) visitation by large numbers of bumblebee queens (Bombus appositus) to Ipomopsis flowers (personal observations; Pleasants and Waser 1985). During 1981 the standing crop nectar volumes in Ipomopsis flowers were much higher than in all but one year ( 1977) of the seven in which they were measured from 1975-1984 (Pleasants and Waser 19 8 5). This apparently permitted bumblebee queens (which cannot normally reach it) to collect the nectar. There was also an unusual asynchrony between the time of peak flowering and the influx of Selasphorus rufus that year (Pleasants and Waser 1985). Together, these observations suggest that there was something unusual about the hummingbird populations at our study site in 1981. It appears that although the number of birds recaptured in 1981 was about that predicted by the regression equation (Fig. 3), the number of new birds was substantially lower than expected (Fig. 4). DiscussionIt is perhaps surprising that we found any significant relationship between flowers and bird abundance, given our crude index; only about 38% of the variation in numbers of banded birds is explained by variation in the four species of flowers. Although the flowers we used are significant resources for the birds, they are not the only species of flowers used by the birds, and they do vary in both temporal availability (Table 1) and in nectar production. The deviations from the regression lines may reflect fluctuations in background population levels of the birds, or in the reliability of our estimates of the floral environment from the hummingbird's point of view. As with any result based solely on correlation, and not explored further with experimental studies, we can suggest causative factors but cannot be positive about the definitive interpretation. Another factor that may cloud the correlation between numbers of birds and flowers is the fact that these birds are long-lived but are not resident at the study site year round; they only spend about two to three months of the year there. The rest of the year is spent in migration, or overwintering farther south (probably in southern Mexico). Thus the availability of food resources during the short breeding season may not be the most important factor regulating population size, even though it may affect the numbers of birds using feeders. Floral resources are also not the only food resources used by the hummingbirds. Insects provide a major source of protein as well as replacement of ions that are lost in the urine (Calder and Hiebert 1983). Hummingbird populations may in fact be more constant than indicated by our data. This would be true if the explanation for increased numbers of banded birds is that during years with lower floral abundance, our population of banded birds derives from a larger area. During these years, for example, some birds probably fly farther to the feeders that we use to attract them for banding, and there might not actually be any significant variation in resident population size; we may just be increasing the size of our effective study area by drawing in more distant birds. The fact that the correlation between recaptured birds and number of flowers is not significant (i.e., numbers of recaptured birds are relatively invariant, with the exception of 1988-1989, when flower numbers were very low; Fig. 3), while that between new birds and number of flowers is significant (without the 1981 datum; Fig. 4), supports the hypothesis that there is a core population of resident birds and a variable number of birds from outlying areas that are drawn to feeders during years of low flower abundance. There is circumstantial evidence to support this hypothesis of a relatively constant core population. For example, the numbers of birds using feeders varies seasonally. Use of feeders is highest during the early part of the season, when flowers are not yet available, and decreases as the availability of floral resources increases. At the end of the season, when floral abundance begins to decline, or when flowers are preempted by the more aggressive migrant Rufous Hummingbirds, use of feeders again increases. At times like this individual birds will fly 2-6 km to our feeders (W. Calder, unpubl. data). The negative correlation between numbers of flowers and banded hummingbirds indicates one difficulty with use of artificial feeders to attract birds for population studies: there is potential for competition from natural food sources. In years with higher floral abundance, numbers of banded birds are lower, apparently reflecting decreased use of the feeders by birds with nests or territories at greater distances. Thus, despite a significant effort at capturing and banding birds each year, our results are biased by flower availability. This pattern differs from that observed for nectarivorous honeyeaters by Pyke (1988); he found that despite a consistent seasonal pattern of daily production of nectar-energy per unit area, the seasonal pattern of honeyeater abundance was not consistent between years, and did not correspond to the pattern of nectar-energy production. He concluded that honeyeater abundances must be determined by factors other than local nectar production. However, Baltosser (1989) found that nectar availability was important to the organization of a guild of hummingbirds, and that they responded to reductions in nectar supplies. He also noted an effect of an unusual freeze on the distribution and number of hummingbird nests in one year. In this case, however, the effect was apparently mediated by damage to potential nest sites rather than floral resources. Despite the variation we have seen in numbers of birds, there does not appear to be any particular long-term trend in either numbers of birds or of flowers at our study site, indicating that these populations may be at some type of equilibrium. The pattern we describe also suggests that there has been no negative effect of the banding study on the hummingbird population (contrary to fears raised by Rosenberg and Stejskal 1988 about the effect of banding). It does, however, suggest that banders who rely on resources such as feeders to attract birds for banding should be aware of the interactions between natural food supplies and feeders. AcknowledgmentsThis research was supported by grants from the National Geographic Society (2665, 2824, 3064, 3242 and 3513; WAC), Earthwatch (DWI and NMW), and the National Science Foundation (DEB8102774, BSR83 13522, BSR8905808: NMW; DEB75 15422, DEB7807784, DEB81108387: 0.R. Taylor and DWI), and a summer Research Award from the University of Maryland Graduate Research Board (DWI). The manuscript benefitted from suggestions from A. D. McGuire, C. A. Kearns, W. H. Baltosser, and two anonymous reviewers. Literature CitedBALTOSSER, W. H. 1989. Nectar availability and habitat selection by hummingbirds in Guadalupe Canyon. Wilson Bull. 101:559-578. CALDER, W. A. 1975. Daylength and the hummingbirds' use of time. Auk 92:81-92. CALDER, W. A., 111. 1981. Heat exchange of nesting hummingbirds in the Rocky Mountains. National Geographic Society Research Reports 13:145-169. CALDER, W. A., 111. 1984. How long is a long-term study? Auk 101:893-894. CALDER, W. A., 111. 1985. The comparative biology of longevity and lifetime energetics. Exp. Gerontol. 20:161-170. CALDER, W. A., AND J. BOOSER. 1973. Nocturnal hypothermia of Broad-tailed Hummingbirds during incubation in nature with ecological correlations. Science 180:751-753. CALDER, W. A., AND S. M. HIEBERT. 1983. Nectar feeding, diuresis, and electrolyte replacement of hummingbirds. Physiol. Zool. 56:325-334. CALDER, W. A., S. M. HIEBERT, N. M. WASER, D. W. INOUYE, AND S. J. MILLER. 1983. Site fidelity, longevity, and population dynamics of Broad-tailed Hummingbirds: a ten year study. Oecologia 56: 689-700. GIBBS, H. L., AND P. R. GRANT. 1987. Ecological consequences of an exceptionally strong El Nino event on Darwin's finches. Ecology 68 :173 5- 1746. GRANT, B. R., AND P. R. GRANT. 1989. Natural selection in a population of Darwin's finches. Am. Nat. 133:377-393. INOUYE, D. W. 1980. The terminology of floral larceny. Ecology 61:1251-1253. INOUYE, D. W. 1988. Natural variation in plant and animal populations, and its implications for studies of recovering ecosystems, p. 39-50. In J. Cairns [ed.], Rehabilitating damaged ecosystems. CRC Press, Boston. INOUYE, D. W. 1989. Environmental control of variation in flowering by two Delphinium species in montane Colorado. Am. J. Bot. 76(6) supplement: 108-109. MILLER, S. J., AND D. W. INOUYE. 1983. Roles of the wing whistle in the territorial behaviour of male Broad-tailed Hummingbirds (Selasphorus platycercus). Anim. Behav. 31:689-700. MULVIHILL, R. 5., AND R. C. LEBERMAN. 1987. Bird banding at Powdermill, 1985: with a summary of Ruby-throated Hummingbird banding data. Powdermill Nature Reserve Research Report #46. Carnegie Institute, Pittsburgh. PLEASANTS, J. M., AND N. M. WASER. 1985. Bumblebee foraging at a "hummingbird" flower: reward economics and floral choice. Am. Midl. Nat. I 14:283-291. PYKE, G. H. 1988. Yearly variation in seasonal patterns of honeyeater abundance, flower density and nectar production in heathland near Sydney. Austr. J. Ecol. 13:1-10. ROSENBERG, G. H., AND D. STEJSKAL. 1988. Arizona. Amer. Birds 42:1325. WASER, N. M. 1978. Competition for hummingbird pollination and sequential flowering in two Colorado wildflowers. Ecology 59:934-944. WASER, N. M. 1988. Comparative pollen and dye transfer by pollinators of Delphinium nelsonii. Funct. Ecol. 2:41-48. WASER, N. M., AND D. W. INOUYE 1977. Implications of recaptures of Broad-tailed Hummingbirds banded in Colorado. Auk 94:393-395. WASER, N. M., AND M. V. PRICE. 1983. Pollinator behaviour and natural selection for flower colour in Delphinium nelsonii. Nature 302:422-424. |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}