|
From Animal Behavior, 1983, 31, 689-700
Roles of the Wing Whistle in the Territorial Behaviour of Male
Broad-tailed Hummingbirds (Selasphorus platycercus)
By Sarah J. Miller & David W. Inouye
Department of Zoology, University of Maryland, college Park,
MD 20742, U.S.A.
Abstract
Suggested roles of avian wing sounds in territorial defence have
not been tested experimentally. The non-facultative, shrill wing
whistle of male broad-tailed hummingbirds occurs during aerial displays
used in defence of courting territories. To investigate the roles
of the wing whistle in territorial behaviour, we observed males
before and after being experimentally silenced, and after having
wing whistles restored. Silenced birds intruded in territories more
easily than normal males, suggesting that the wing whistle is a
conspicuous signal advertising the presence of adult, male birds.
Birds were less active and aggressive in territory defence after
silencing than they were either before silencing or after restoration
of the wing whistle. Silenced birds tended to lose their territories
to rival hummingbirds more readily than non-silenced birds. We propose
that silenced birds defend their territories less effectively because
they cannot communicate threat and because they lack sensory feedback
that normally enhances a bird's aggressiveness. Results of this
study suggest that the wing whistle of male broad-tailed hummingbirds
is important in maintenance of courting territories and hence in
reproductive success.
Sound production in animals has evolved in many cases to enhance
communication among individuals. It has long been thought that some
bird songs function in territory advertisement (Thorpe 1961; Armstrong
1963), and recent experimental evidence supports this theory (Peck
1972; Smith 1976; Krebs 1977; Smith 1979). There has been less investigation
of the potential territorial roles of mechanically produced avian
sounds. It has been suggested that wing sounds in spruce grouse
(MacDonald 1968), flappet larks (Wickler 1967; Payne 1973; Bertram
1977), and woodcocks (Sheldon 1967) probably function in territorial
advertisement, as do tail feathers of snipe (Jellis 1977). Wing
rattling in todies occurs during territory defence (Kepler 1977),
and drumming in woodpeckers may facilitate territory defence (Winkler
& Short 1978). Direct evidence has not been provided to test the
behavioural roles of any of these sounds.
Male hummingbirds (Trochilidae) are typically territorial, defending
mating and feeding territories during the breeding season (Wolf
& Stiles 1970), and feeding territories at other times (Pitelka
1942). Males of a number of hummingbird species reportedly produce
wing sounds in flight. These species include black-chinned hummingbirds
(Archilocus alexandri), Costa's hummingbirds (Calypte
costa), broad-billed hummingbirds (Cynanthus latirostris)
(Bent 1964), Lucifer hummingbirds (Calothorax lucifer), and
violet sabrewings (Campylopterus hemileucurus) (Wagner 1966).
Wing sounds are also produced by members of the genus Selasphorus,
including three temperate zone species, the broad-tailed hummingbird
(S. platycercus), the rufous hummingbird (S. rufus),
and the Allen's hummingbird (S. sasin), and by at least one
tropical species, the scintillant hummingbird (S. scintilla).
The wing sound of S. rufus is thought to aid in courtship
of female conspecifics and intimidation of male conspecifics (Sprot
1927). Pitelka (1951) suggested that the wing sound of S. sasin
announces the presence of males during circuit flights over their
territories.
During rapid flight male broad-tailed hummingbirds (S. platycercus)
produce a particularly shrill, buzzing whistle as air rushes through
slots created by the tapered tips of the ninth and tenth primary
wing feathers (Fig. 1) (Ridgeway
1877; Woodbury & Sugden 1938; Wagner 1948; Armstrong 1963). It is
uncertain whether the wing whistle is produced all year round or
primarily during the breeding season (see Henshaw in Ridgeway 1892,
and Wagner 1948 for opinions). The sound resembles the buzz of a
cicada (Cicadidae) (Bailey 1928). The sound is probably perceptible
at 50 m by males, and at more than 75 m by female conspecifics (Wagner
1948). Ridgeway (1877) wrote that dives and chases by males, "accompanied
as they were by the shrill piercing noise, invariably put to flight
all birds assaulted". Linsdale (1938) documented an especially
loud buzz occurring in flight after a territorial chase. According
to Wagner (1948, 1966), the wing whistle, which is obvious during
territorial display flights, may be used in mate attraction and
courtship.
The purpose of this study was to investigate the suggested roles
of the wing whistle of male broad-tailed hummingbirds in territory
defence. If the wing whistle functions as a social signal, its removal
should result in behavioural changes in manipulated birds as well
as the birds around them. Specifically, if the wing whistle is an
aggressive signal, wing-silenced birds might be less effective in
territory defence, substitute other aggressive behaviour for the
missing whistle, or switch to less expensive defences if energy
became limited. Flexibility in territorial behaviour has been documented
as a response to variables such as resource quality (e.g. Ewald
& Carpenter 1978). Since changes in an aggressive signal should
affect the behaviour of the signal's receivers, we also predicted
that conspecific males should be more aggressive and effective than
normal at intruding into territories of silenced males. To test
these predictions, territorial, male broad-tailed hummingbirds were
observed before and after experimental silencing of the wing whistle,
and interactions of these birds with conspecific males were analysed.
Methods
Study Site
We observed breeding broad-tailed hummingbirds between 29 May and
7 July 1981 at the Rocky Mountain Biological Laboratory, located
at an elevation of 2900 m in the Elk Mountains of Central Colorado.
The laboratory is situated in a steep valley dominated by subalpine
meadow that is interrupted by patches of willows (Salix sp.).
Adult males defended courting territories for approximately 6 weeks,
from early June to mid-July. Post-breeding rufous hummingbirds began
to migrate south through the study site in early July. An estimated
13 of 62 (21%) male broad-tailed hummingbirds at the study site
were territorial, while the remainder were non-territorial and presumably
competed for territory sites. We found 13 courting territories of
male broad-tailed hummingbirds in the study area; they surrounded
and enclosed willow patches and varied in size from 1800 square
metres to 8000 square metres (X = 4343.9 square metres, N
= 10). We estimated territory sizes from chase distances, location
of intruders that elicited chase responses, and location of circuit
flights. Willows, typically 1 m in height, covered 60-70% of the
area of any given territory and served as perches.
Courtship and probably copulation occurred within territories, but
nesting did not. Nectar sources within territories were rare except
for willow blossoms, which were used by hummingbirds for 2 weeks
in early June (personal observation). We placed a single feeder
containing 30% sucrose (by volume) solution at the approximate centre
of each territory to control for variation in food availability
to territory owners. Territories were established before we distributed
feeders, hence territories would have been defended without the
supplemental feeding. All observations were made at least 1-1.5
days after birds had established territories, to control for the
amount of time required for new owners to equilibrate in terms of
defence costs (Copenhaver & Ewald 1980).
Capture and Mark Techniques
Most hummingbirds were captured using traps; butterfly nets and
hummingbird mist nets were also used (capture permit 20753-A, first
author, and 20753, second author). Traps were constructed by sewing
fibreglass insect screening to a wire frame 0.6 m long and 0.4 m
in diameter. A feeder was hung inside and a trap door could be closed
using guy lines operated by a person hidden in the willows several
metres behind the trap.
Two methods were used to mark captured birds. Coloured flagging
was glued to the hummingbird's back using Duco cement (see methods
in Baltosser 1978). Alternatively, Hyplar acrylic paints of similar
colours were used to place a dot of colour on the bird's back or
to paint the three central rectrices. Markings were easily visible
with binoculars and frequently with the naked eye. Neither capturing
nor marking had apparent effects on the territorial behaviour of
most birds.
Observations
We chose to observe seven territory sites that were centrally located
in the study area, with two of the sites separated by about 50 m
and 250 m, respectively, from the others, and five of the sites
clumped with shared borders. These seven sites offered high fequencies
of behavioural interactions. Five of the seven sites were occupied
by one individual throughout the entire breeding season, whereas
a turnover of owners occurred at the others. We observed eight territorial
males, one of which served first as a control and later as an experimental
subject.
Times of observation were typically 0700 to 1200 hours because territorial
activity was high and relatively constant during morning hours.
Observers sat approximately 5 m from territory boundaries and recorded
frequencies and durations of behavioural events important in territorial
defence. Events observed included chases, vocal chatter, circuit
flights (flights around a territory within its boundaries, sensu
Pitelka (1951), occurring either in response to males heard outside
the territory or when no environmental stimulus was evident), pendulum
dives (vertical, U-shaped dives approximately 30 m in height (Bene
1947; Wagner 1948)), shallow dives directed at conspecifics, and
the presence of intruding or passing hummingbirds. These flights
and chattering are frequent and important in territory defence in
many temperate zone hummingbirds (Pitelka 1951; Lyon 1973; Ewald
& Carpenter 1978).
Experiments
The male birds used in our experiments were each observed for 2
days before treatment to silence the wing whistle. To fully silence
the sound, we applied a thin film of Bond 527 quick-drying glue
(Bond Adhesives Co.) with a toothpick to the narrowed portions of
the ninth and tenth primary wing feathers (i.e. the distal 7 mm
for the thenth, and distal 3 mm for the ninth). Glue weighed less
than 0.024% of a bird's weight (mean bird weight = 3.58 g, N
= 4; glue dry weight = 0.00085 g measured by a Mettler H20 balance).
According to formulas presented by Feinsinger et al. (1979), we
estimated that the wing disc loading (mass of the bird/area swept
out by the wing) was thereby increased by 0.02%, and the cost of
hovering flight was increased by 0.01%. The cost of forward flight
is probably less than that of hovering flight (Greenewalt 1975).
Experimental birds probably could compensate for the slight increase
in flight costs by ingesting more food from feeders available at
territory sites. Silenced birds did not preen more often than usual
or exhibit any behavioural changes other than those recorded in
interactions with conspecifics.
We observed the experimental birds again after silencing, and thus
each bird served as its own control. Silenced birds on their territories
were just as easily observed as normal, non-silenced birds, and
hence observations were not biased against silenced birds. Five
birds were each observed for the 2 or 3 days following silencing.
Two others were observed until they abandoned their territories
on the tenth and fourteenth days, respectively, after silencing.
Sample sizes of experimental silencing trials were small because
of (1) the limited number of males with territories having sufficient
behavioural interactions, and (2) the difficulties with recapturing
techniques at the start of the season.
The wing whistles of three birds were restored by removing the glue
with acetone; glued portions were dipped in acetone and swabbed
with acetone-soaked cotton swabs, during which proper ventilation
was used to prevent fumes from affecting the birds. Feathers appeared
completely normal after treatment with acetone.
Two sham-glued birds, with glue placed on equivalent surface areas
of non-narrowed portions of primary wing feathers, served as controls
for the presence of glue on the wings. Additionally, the acetone
treatment, a procedure requiring manipulations similar to those
of the gluing procedure, caused none of the behavioural changes
observed in glued birds.
Observations of control birds were paired with appropriate experimental
birds to control for daily and seasonal variations in territorial
behaviour. Territorial activity varied among days because of weather
(e.g. on windy days birds were less active), and varied seasonally
with amount of courtship activity and with number of conspecific
and heterospecific hummingbirds present at the study site. One control
bird, which had been captured and marked, served as the control
for one of the seven silencing trials. This bird's territory was
located adjacent to that of the experimental bird. A second control
bird was captured and marked, first silenced for 6 days, then treated
with acetone to restore its wing whistle. This bird served as the
control for the other six trials, and defended a territory approximately
250 m from those of the experimental birds.
Analysis
Tapes of wing whistles and vocalizations of several territorial
males were made using a Uher 4000 Report IC Tape Recorder with a
Uher M517 microphone mounted in a parabolic reflector. Durations
and frequencies of territorial events were analysed using a randomization
test to compare pre- and post-treatment behaviour in experimental
birds. Control data were not subjected to statistical analysis since
the same bird was observed as a control for six of the seven trials.
However, mean control data for the seven trials are portrayed graphically
with mean experimental data. Percentage data were arc-sine transformed.
Behavioural events with frequencies less than or equal to 2/h were
not included in analysis of data with percentages; this involved
observations during less than 10% of the total hours.
Results
Territory Defence
When the wing whistles of territorial males were experimentally
silenced, their territorial behaviour altered. This was not the
case with the control birds with which experimental birds were paired
in time (but not in a statistical sense). Experimental birds chased
fewer intruding males after being silenced than before (P
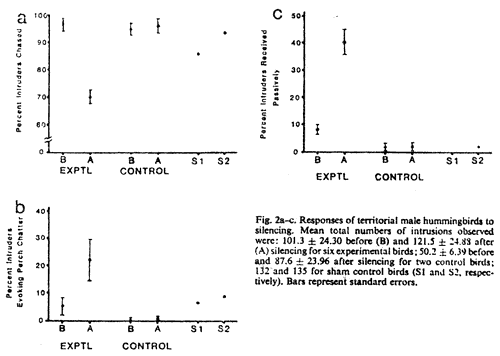
= 0.03, N = 6 birds, Fig. 2a).
They perch-chattered (i.e. remained perched while chattering, instead
of initiating a chase within 1 s) at more intruders after being
silenced (P = 0.03, N = 6 birds, Fig.
2b, often allowing intruders to fly to them, hover over them,
or direct shallow dives at them. Twice, we observed adult, male
conspecifics directing pendulum dives at silenced territury owners,
while we observed none directed at normal, non-silenced owners.
A territory owner was considered "passive" if it remained
perched while allowing any of these types of intruder behaviour,
or if it was chased by an intruder. More intruders were received
passively by territory owners (i.e. owners were passive during more
territory intrusions) after the owners were silenced than before
(P = 0.016, N = 6 birds, Fig.
2c).
Territorial chases by silenced birds appeared to be slower than
those of non-silenced and sham-glued birds, although flight speeds
were not measured. Fewer territorial chases by silenced birds were
successful relative to those of non-silenced birds (P = 0.03,
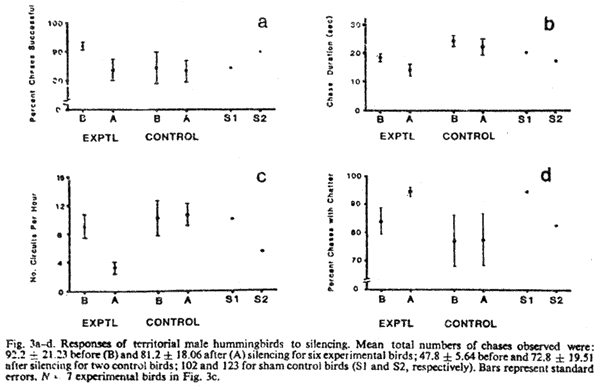
N = 6 birds, Fig. 3a);
an unsuccessful chase was one in which the intruder hovered, chased,
or dived at the silenced owner rather than leaving the territory
readily. Means for the control birds in Fig.
3a are low because the bird that served as a control for six
of the silencing trials directed most of its chases at a neighbouring
male during boundary disputes. The neighbouring male apparently
defended portions of the control bird's territory, and was therefore
unusual in that it did not leave the control bird's territory readily.
Chases were shorter in duration (P = 0.03, N = 6 birds,
Fig. 3b) and in distance after
birds were silenced; chases frequently terminated within territorial
boundaries rather than extending beyond them as chases did before
birds were silenced. Territory owners performed fewer circuit flights
after silencing than before (P = 0.016, N = 7 birds,
Fig. 3c). The number of male intruders
per hour did not alter significantly after territory owners were
silenced (X before = 9.3, X after = 9.2, P
> 0.05, N = 7 birds)>
To investigate compensation for lack of the wing whistle by use
of chatter during flight, frequencies of chatter vocalizations were
measured. More chases involved chatter by territory owners after
birds were silenced than before (P = 0.03, N = 6 birds,
Fig. 3d) More circuit flights
involved chatter in four of six birds after silencing than before
(X before = 15.5, X after = 28.1, P > 0.05,
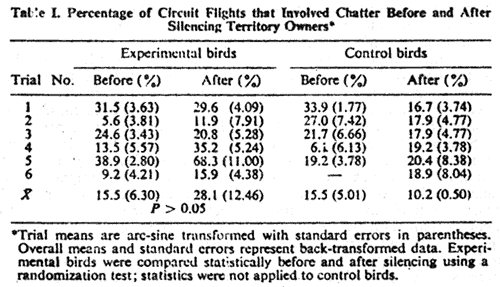
N = 6 birds, Table I).
Durations of chatter during chases and circuit flights were not
recorded.
Territory sizes decreased by an average of 36.4% (N = 8 territories,
measured before and 2 days after owners were silenced), presumably
as a result of delayed chase responses and shorter chase distances
of territory owners following silencing. Two territory owners were
observed for a week after being silenced to test the possibility
that birds might eventually return to normal behaviour with normal
territory sizes. Data for neither bird showed any significant return
to normal behaviour in the 10 variables measured.
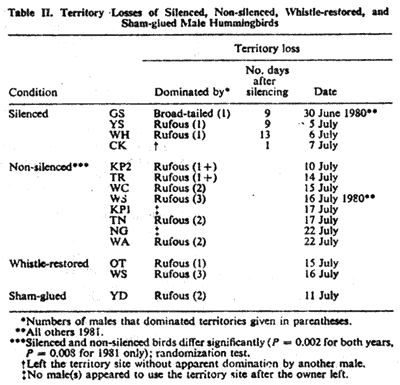
Data on territory losses to rival males, mostly migrant male rufous
hummingbirds, suggest that silenced broad-tailed hummingbirds tended
to lose their territories more readily than non-silenced conspecifics.
Four silenced birds left or lost their territories to rival males
by early July; two of these were dominated by single, migrant rufous
hummingbirds and one by a single conspecific (Table
II). Seven of eight non-silenced birds, and both birds with
wing whistles restored, did not abandon their territories until
14 July or later, and then mostly because they were dominated aggressively
by large numbers of rufous hummingbirds (Miller, in press a). Dates
of territory loss (or abandonment) differ by a randomization test
for silenced and non-silenced birds, both for 1981 alone (P
= 0.008) and for 1980 and 1981 together (P = 0.002). Whistle-restored
birds and a sham-glued bird fall within the range of dates for non-sioenced
birds. Birds not represented in Table
II had died or left the study area because of experimental manipulation.
Non-silenced birds apparently maintained territories later in the
season than silenced birds and resisted domination by single rival
males. More complete data on the relative length of territory ownership
for silenced and non-silenced birds are not available.
Four birds were observed immediately after silencing to determine
whether the passiveness of silenced birds (1) was a delayed response
to intruder behaviour, or (2) was an immediate response to loss
of the wing whistle. The first alternative is correct if freshly-silenced
birds used normal "beeline" chases (direct, rapid chases
begun as soon as intruders entered territories) during initial encounters,
but later used passive, delayed responses. Two birds used beeline
chases when first silenced to exclude the first four intruders and
the first intruder, respectively. All subsequent chases were more
passive. Two birds did not use beeline chases at any time following
silencing; their behaviour supports the second alternative. No definite
conclusion may be drawn.
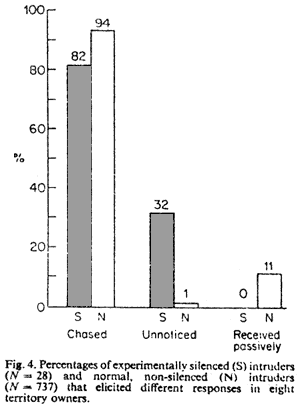
Territory Intrusion
Territory owners responded to experimentally silenced, intruding
male conspecifics differently than to normal, non-silenced intruders.
We observed two different owners directing pendulum dives at a silenced
intruder, whereas we never observed owners directing pendulum dives
at non-silenced, adult intruders. Silenced intruders, more than
normal intruders, went unnoticed altogether or were unnoticed initially
upon entry before being chased out by territory owners (Fig.
4). Flight patterns of silenced intruders that might have caused
them to go unnoticed, e.g. flying low in the vegetation, were not
observed. Although both silenced and normal intruders were chased
off territories eventually, territory owners initially behaved passively
toward normal intruders more often than toward silenced intruders
(Fig. 4).
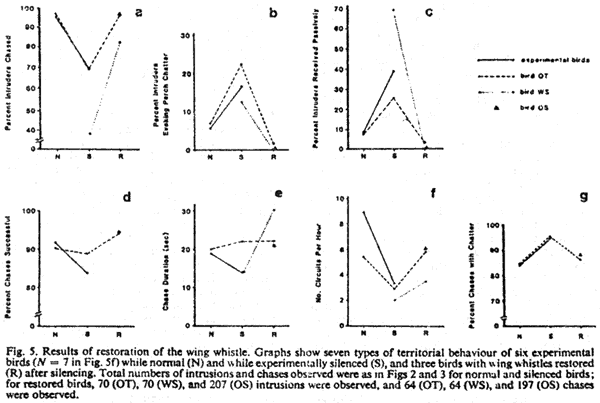
Restoration of the Wing Whistle
Male hummingbirds exhibited a return to normal territorial behaviour
accompanying the return of the wing whistle. For bird OT, as shown
in Fig. 5, most variables that
altered significantly in all birds as a result of silencing returned
after whistle restoration to almost exactly the same levels as before
silencing. The only variable that did not alter as a result of silencing
for bird OT was chase duration (Fig.
5e); nevertheless, it was the same after whistle restoration
as before silencing. For two other birds with restored wing whistles,
no observations were made before silencing and hence we cannot assert
that they returned to presilenced levels of behaviour.
The three whistle-restored birds did not differ from the six (seven
for Fig. 5f) non-silenced birds
for variables in Figs 5a, 5e, and
5f (P > 0.05; randomization test). They did differ for
variables in Figs 5b and 5c (P
= 0.02; randomization test) but not in the predicted direction;
that is, whistle-restored birds were even less passive toward intruders
(less like silenced birds) than were non-silenced birds.
Sham Controls
Two sham-glued birds functioned as controls for the presence of
glue on the primary wing feathers. Sham bird 2 (S2), with glue placed
on the tips of the seventh and eighth primaries, produced an essentially
normal wing whistle. Sham bird 1 (S1), with glue placed over equivalent
surface areas on proximal portions of the ninth and tenth primaries,
produced no wing whistle for two days following treatment. Within
a week the whistle was produced again, but it varied in volume (from
about 75% to 100% of that of normal whistles) during the remainder
of the study. During several types of flights in which other birds
produced whistles of low folume, bird S1 was silent.
It is difficult to interpret data for the sham-glued birds because
of the small sample size. For several behavioural variables, the
sham-glued birds were normal relative to untreated birds (Figs
2a, 2b, 2c, 3b), whereas for other variables, at least one sham-glued
bird behaved more like silenced birds (*Figs 3a, 3c, 3d). The frequency
of circuit flights for sham bird S2 actually falls above the highest
values for silenced birds, and below values for six of the eight
non-silenced birds (Fig. 3c).
Without a larger sample size, we consider sham-glued birds to have
behaved normally in terms of frequency of circuit flights. However,
sham bird S1 performed fewer chases successfully (Fig.
3a) and more chases with chatter (Fig.
3d) than untreated birds. The values of these two variables
may be interpreted in light of the fact that bird S1 produced a
wing whistle of subnormal volume at times; this might cause some
compensation with chatter during flight, and might cause some intruders
to turn back while being chased and investigate the territory owner
(bird S1) (see Discussion).
Hence, variations from predicted values in data for sham-glued birds
might be explained by slight auditory consequences of the sham-gluing
treatment, rather than the simple presence of glue on the primaries.
In the absence of stronger, more complete controls for the presence
of glue, we cautiously interpret the present data to indicate that
gluing of the primaries did not itself cause behavioural changes
in experimental birds.
Wing Whistle Audibility
The wing whistle was often audible to the human ear from distances
of 75-100 m. Evidence that the wing whistle was audible to conspecific
birds comes from observations of territorial males; we noticed that
other males that we heard faintly in the distance elicited vocal
chatter in the territorial males. Apparently the whistle was at
least as perceptible to these males as to us.
We also observed conspecific responses to the wing whistle at closer
ranges. Once, while a female was perched under a porch roof, a male
performed a pendulum dive that was directly above her, but was visually
obstructed by the roof. The female flinched as the male reached
the bottom, loudest portion of the dive. Numerous times, when we
heard a male aproaching a hummingbird feeder from around the corner
of a cabin, we watched feeding females fly rapidly in the opposite
direction, and we observed feeding males quickly approach the oncoming
male.
Discussion
The wing whistle is a loud, shrill sound that is conspicuous during
nearly all flights of male broad-tailed hummingbirds. As such, the
wing whistle announces the sex, adult status, and presence of a
male. It is probably an important means of communication among males
who presumably compete for courting sites and for mates.
The whistle renders males conspicuous to rival males. Because silenced
intruders fail to advertise their presence, they go unnoticed by
territory owners more often than do non-silenced intruders (Fig.
4). Intruding silenced birds are therefore probably more successful
at acquiring resources (although silenced and normal intruders alike
are eventually chased from territories; Fig.
4). Kodric-Brown & Brown (1978) proposed that the wing sound
of adult male rufous hummingbirds (S. rufus) reduces their
success at intruding relative to conspecific females and juveniles.
Data from Ewald & Rohwer (1980) for Anna's hummingbirds (Calypte
anna) support an alternative: the brighter coloration of adult
hummingbirds reduces their intrusion success. Our data from wing-silenced
S. platycercus indicate that wing sounds of hummingbirds
affect the success of territory intrusions.
The wing whistle probably plays an important role in advertising
the presence of territorial males at particular sites. Circuit flights
of hummingbirds may serve this function (Wagner 1948; Pitelka 1951),
and in male broad-tailed hummingbirds other audible, territorial
flights such as dives and chases probably do also. If the whistle
during flights of territorial hummingbirds advertises presence and
deters potential intruders, then silenced territory owners performing
relatively few such (silent) flights might be expected to have relatively
high territory intrusion rates. The number of intrusions per hour
per territory did not change after we silenced territory owners;
however, a better measure would have been intrusion rate/unit area
since territory sizes decreased after owners were silenced. It is
likely, in light of evidence that territory size often reflects
changes in intrusion pressure (Myers et al. 1979; Ewald et al. 1980),
that higher intrusion rates were hidden by obervation of intruders
over smaller areas.
Role in Aggression
Results of the present study indicate that the wing whistle of
male broad-tailed hummingbirds is important in the aggressive defence
of territories. We suggest that experimentally-silenced birds may
be less effective in territory defence than normal birds because
(1) silenced birds cannot communicate threat during territorial
displays, and (2) they act less aggressively because their normal
behaviour patterns are disrupted by the sudden loss of the shrill
sound that typically accompanies flight.
Threat. Territorial hummingbirds are notoriously
aggressive, and in species such as broad-tailed hummingbirds which
lack song, display flights (such as chases and dives) may intimidate
territory intruders (Pitelka 1942). The wing whistle is most noticeable
during these territorial displays and apparently enhances the communication
of threat.
Specifically, our data show that chases by silenced territorial
males are less successful than those of normal males in deterring
intruders (Fig. 3a), and conspecific
responses to a male's wing whistle indicate it is a threat. Also,
silenced territorial birds appear to substitute chatter for wing
whistle threat signals. In physical structure, the chatter vocalization
is characteristic of aggressive sounds (i.e. it is harsh, covering
a wide-frequency band, and is of relatively low frequency; Morton
1977).
Territorial encounters with silenced birds indicate that they are
not perceived as threatening. Pendulum dives, which are normally
directed toward any bird except another conspecific, adult male,
are directed toward silenced, adult males, and shallow dives, normally
directed only toward rival males, are not directed toward silenced
males. Also, the silent flight of silenced intruders is incongruous
with their flight behaviour (which is not the unobtrusive flight
behaviour characteristic of silent females). Territory owners consistently
investigate and chase thses non-threatening, abnormal intruders
more than non-silenced intruders (Fig.
4).
Results of a sound analysis suggest that the wing whistle of male
broad-tailed hummingbirds varies in sound frequency and volume with
the level of aggressive motivation of the bird (Miller, in press
b). This graded threat may be advantageous to a territorial hummingbird
because energetically, a warning costs less than an attack, and
also allows males to settle conflicts on the basis of differing
motivations (van Rhijn 1980).
Enhancement of aggressive state. The wing whistle is undoubtedly
extremely loud to the bird producing it. The sound itself, or vibrations
associated with it, may create sensory feedback, enhancing the aggressive
motivational state of the bird. If so, the sudden loss of this sound
would disrupt the normal behaviour of the bird because the bird
may no longer perceive itself as a normal male. Our data suggest
the males are less aggressive after silencing, implying that the
wing whistle enhances the aggressive state of male.
Experimentally silenced territory owners often perch low in the
vegetation, have difficulty expelling intruders (Fig.
3a), and ultimately tend to lose their territories to rival
males (Table II). Fewer intruders
are chased by silenced owners (Fig.
2a), and individual chases are less intense because of delayed
chase responses (Fig. 2c), shorter
chase durations (Fig. 3b), and
slower, less direct flight. Fast, direct flight is generally a threatening
behaviour (Brown 1975). Chases of non-silenced birds but not silenced
birds typically extend far beyond territory boundaries, a behaviour
that in hummingbirds may deter intruders by indicating aggressive
ability and ownership (Copenhaver & Ewald 1980).
DIsruption of normal aggressive behaviour is also implied by the
difficulty of silenced birds in establishing new territories. We
saw no silenced, territorial males (of 11) usurp territories or
establish themselves on new sites, whereas of approximately 50 non-silenced
males, at least 15 established new sites during the course of the
summer in 1981, and at least five usurped sites. Silenced males
may not have attempted territory establishment because of their
disrupted aggressive state, or they may have made unsuccessful attempts,
but in either case their aggressive abilities apparently were reduced.
All the data thus imply that the wing whistle enhances territorial
aggression.
An alternative explanation for the changes in territorial behaviour
after birds are silenced might be that experimental birds hide their
abnormal silence by switching to defences that involve less flight.
It is difficult to disprove either explanation with the data at
hand, but comparison of the present study with similar studies (see
below) provides support for our suggestion that loss of the wing
whistle causes loss of aggressive motivation.
Evolution of the Wing Whistle
Sounds occurring in the repertoire of a species presumably have
been selectively favoured because they increase the fitness of the
producer. Since broad-tailed hummingbirds defend feeding territories
during the non-breeding season (DesGranges & Grant 1980), and defend
courtship territories used in their promiscuous, competitive mating
system (Wolf & Stiles 1970; this study), both the feeding and mating
systems could provide selection for an effective aggressive signal.
Wagner (1966) has suggested that the wing whistle is under sexual
selection for mate attraction and stimulation. Production of sensory
feedback that enhances a hummingbird's aggressive state may be a
secondarily-derived advantage correlating with the type of social
signal that evolved. Selection for a social signal used during the
breeding season may be particularly intense because of the interspecific
dominance of migrant rufous hummingbirds, whose exclusion of broad-tailed
hummingbirds from territories in July (Miller, in press a) may place
a constraint on the breeding season of broad-tailed hummingbirds.
This constraint seems likely, since a socially subordinate species
usually narrows its niche when sympatric with a socially dominant
species (Morse 1974). The occurrance of mechanically-produced wing
sounds among hummingbirds does not correlate either with migratory
habits or with the use of high-elevation habitats. However, the
open habitat of broad-tailed hummingbirds may be important in selecting
for the use of conspicuous aerial displays enhances by acoustic
signals.
Relevance to Similar Studies
Results of the experimental removal of the wing whistle of male
broad-tailed hummingbirds resemble those of other studies involving
the removal of socially significant visual or acoustic signals from
male animals. Removal of antlers of red deer (Lincoln 1972), of
epaulet coloration of red-winged blackbirds (Peek 1972; Smith 1972),
of dark throat plumage of Harris sparrows (Rohwer 1977), and of
face masks of yellowthroats (Lewis 1972) reduced their success in
social aggressive encounters. However, in these cases, unlike in
the present study, experimental males spent more time and energy
than normal in intrasexual behavioural conflicts in an attempt to
maintain territories or social dominance. The increased aggressive
behaviour apparently was a response to the bold behaviour of surrounding
individuals.
The signals investigated in the above studies appear to have varying
importance in competitive encounters. For instance, experimental
Harris sparrows compensated for loss of the ability to signal dominance
by using other aggressive behaviour; experimental red-winged blackbirds
compensated to some degree, but they often experienced territory
loss; and experimental red deer stags soon adopted submissive behaviour
and avoided conflicts even though in initial conflicts they were
successful in maintaining dominance. Experimental hummingbirds in
the present study fall even farther along this continuum because
they adopted non-aggressive behaviour immediately after they could
not signal aggressive intent.
We suggest that the aggressive motivation of red deer and broad-tailed
hummingbirds were the most disrupted, red-winged blackbirds less
so, and Harris sparrows the least disrupted by releaser removal.
The differences in the awareness of signals are reflected in the
ranking of disruption of aggressive motivation.
There is evidence from experiments with muted red-winged blackbirds
that may be contrary to our theory. If song creates sensory feedback
enhancing aggressiveness, then muting should cause reduced aggressiveness.
Peek (1972) and Smith (1976) found that males with muted song continued
to display visually and apparently defended territories vigorously
and effectively. However, in Smith's (1979) study, muted birds (N
= 3) experienced higher intrusion rates and lost their territories,
suggesting disruption of aggressive motivation. Also, although 3
of 12 muted males in Peek's (1972) study were aggressive enough
to regain territories that they had lost after they were muted,
these birds had little competition in doing so. If muted red-winged
blackbirds do not lose aggressive motivation, it might be because
either (1) their muted vocal systems still produce some sound (Smith
1976), or (2) the threat display is still partly functional because
birds can still display visually by spreading their wings. In none
of the studies of muted red-winged blackbirds were the researchers
investigating loss of aggressive motivation, hence more data are
needed to test our theory of feedback enhancement of motivation.
The above studies of other animal species involving the removal
of releasers used in social aggression generally support the hypothesized
functions of the wing whistle in communicating threat and enhancing
the aggressive state by physical feedback during territory defence
by male broad-tailed hummingbirds. Results of the present study
also suggest that the wing whistle advertises the presence of an
adult, male hummingbird. This conspicuous acoustic signal, by mediating
intrasexual conflicts for mates that visit courting territories
during the breeding season, may enhance reproductive success in
male broad-tailed hummingbirds. Further investigation is needed
to test potential roles of the wing whistle in courtship and in
winter territoriality.
Acknowledgments
This paper is part of a Master's thesis submitted to the University
of Maryland by the senior author. We gratefully acknowledge the
many forms of help provided by Wolfgang M. Schleidt and Eugene S.
Morton throughout the study. Allen W. Smeltzer designed the hummingbird
trap and offered much encouragement; he and Rebecca Craft provided
invaluable field assistance. Statistical advice was patiently given
by Svata M. Louda, Estelle Russek, and Stephen G. Vail. William
A. Calder III and Nicholas M. Waser helped band many of the birds
used in this study, and they offered ideas and equipment for field
work. Critical reading of drafts of various manuscripts was provided
by Wolfgang M. Schleidt, Eugene S. Morton, Robert S. Fritz, Paul
W. Ewald, and many others. This research was supported by grants
to S.J.M. from the Frank M. Chapman Memorial Fund, the Harris Foundation,
and Sigma Xi, the Scientific Research Society, and NSF Grant DEB
78-07784 to D.W.I.
[Note: the references for this paper were illegible on my photocopy.
If you need them, please ask your librarian to obtain a copy of
the journal, or write the author for a reprint. -LC]
|

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}